>>> Click here to access this episode of the Syllab Podcast on Spotify <<<
a) Purposes and functions of the musculoskeletal system
We all have a fairly good high-level mental representation of our skeleton and muscles, though certainly not in the details, for the simple reason that the outlines of both are apparent to the naked eye and we get to directly experience various of their functions, specifically the ability to hold ourselves upright and move around. Muscles and skeleton form a system called the musculoskeletal system for two main reasons: the first is that they are physically linked and the second is that some of the functions they perform require the presence of both; all evident signs they would have evolved alongside each other.
Functions. Evolution. The usual terms I bring up when it comes to biological organisms. And to help us understand why some functions evolve, it pays to take one step back and consider purpose, in the sense of the type of benefit each function brings to the organism from an evolutionary fitness perspective. In the case of the musculoskeletal system, the main functions could be grouped into four: movement, stability, organ protection, and form.
It is pretty straightforward to ascertain the actions permitted by movement: escape predators, find mates, and obtain food and therefore nutrients required by the organism to power itself and maintain homeostasis. In order, those have a role in terms of survival, reproduction, and survival again. With regards to stability, it is mainly a matter of avoiding falls and injuries, a role accomplished by proprioception as part of our vestibular system and performed under the unrelenting supervision of the central nervous system, thus it comes down to survival. Organ protection is also about improving the odds of survival by shielding key vital organs from potential external damages. This is why our lungs are enclosed within a rib cage and our brain has its own reinforced apartments inside our skull or cranium. Form is slightly less obvious, it allows for spatial awareness by our nervous system, which is both useful to maintain stability and for movement, and it helps ensure internal coordination by making possible the systematic and consistent location of any organ within a given organism without which cell differentiation would be of little use and gene regulation would be unfeasible at the level of large complex entities (refer to Series 1 Section 3.c for discussion on gene regulation and S1 Section 5.c for cell regulation).
The musculoskeletal system also renders other services to the organisms, but I would qualify them of secondary functions as they do not fit in the same logic of driving in a specific evolutionary direction, rather they appear to be exercises in optimization; making do with what is already there. The main secondary functions worth mentioning in this essay, bearing in mind this is not an exercise in exhaustive description of the human body, are the production of blood cells and the storage of minerals.
The bone marrow is a tissue found in our long bones, parts of our skeleton, it plays a key role in our immune and circulatory systems, topics covered in Chapter 5 and Chapter 1 respectively, by producing and housing white blood cells, red blood cells and the platelets that can prevent haemorrhage from punctured blood vessels by forming blood clots.
As for minerals’ storage, the main elements involved are calcium and phosphorus. In Section 2.f we have seen the latter goes into the make-up of our energy currency, the ATP macromolecule, and our genetic material, RNA and DNA, but it is also involved in the regulation of blood pH and in the activation of certain enzymes. The kidneys are the main organ ensuring this regulation and the phosphorus is deposited in the bones alongside calcium in the form of hydroxyapatite crystals which account, in a slightly modified form known as bone mineral, for nearly 70% of the total weight of our bones. Besides ensuring bone strength, calcium is a key element in various parts of our organism, whether it is blood vessel contraction and dilation, hormonal secretion or muscle contraction, something we’ll look into in a brief moment, though not solely from the standpoint of calcium’s role.
b) Muscles contractions and locomotion
Going back to the primary function of movement, the term is meant to capture both the change in the position of parts of our body in relation to the rest of the organism like the raising of an arm as well as the translation in space of the entire body, what we call locomotion. I will deal with locomotion in the next section because its performance requires the skeleton, in addition to the muscles, and it hasn’t yet been properly introduced.
The mechanism underlying movement is muscle contraction. In the human organism, muscles come in three different types. The first is the smooth muscle cells lining the walls of organs such as the stomach and blood vessels, they do not join together to form fibres and their contraction or dilation is initiated in the muscles themselves though with modulation ensured by the autonomic nervous system but without mediation by nerves. The second is the cardiac muscle which is also regulated by the autonomous nervous system rather than the central one but, there, strips of muscle cells join together to form long fibres. The third type of muscle, the one we’ll focus on going forward is called skeletal muscle, it is the type attached to the skeleton and permitting movement. It is innervated since its contraction is directed by the central nervous system. We will discuss the autonomous and central nervous systems in Chapter 6.
Like the cardiac muscle, the tissue of skeletal muscles consists of fibres formed by the fusion of cells, a process called myogenesis during which the various nuclei and mitochondria are preserved. This is a simple enough description but it won’t do in terms of explanation. To truly understand how muscles contract we need to scrutinize the makeup of those fibres at the macromolecular level and the biochemistry taking place there. The functional unit of reference in fibrous or “striated” muscle tissues is the sarcomere, the element from which the striated appearance originates. These units actually consist of alternating elongated proteins, mostly myosin and actin, which slide past each other and when they do so, since the actin filaments are anchored into Z-lines separating series of sarcomeres, those Z-lines get closer to each other, thus creating an overall contraction of the muscle fibre. Figure 2 below provides a diagrammatic illustration of this organic set up.
Figure 2: Sarcomere contraction
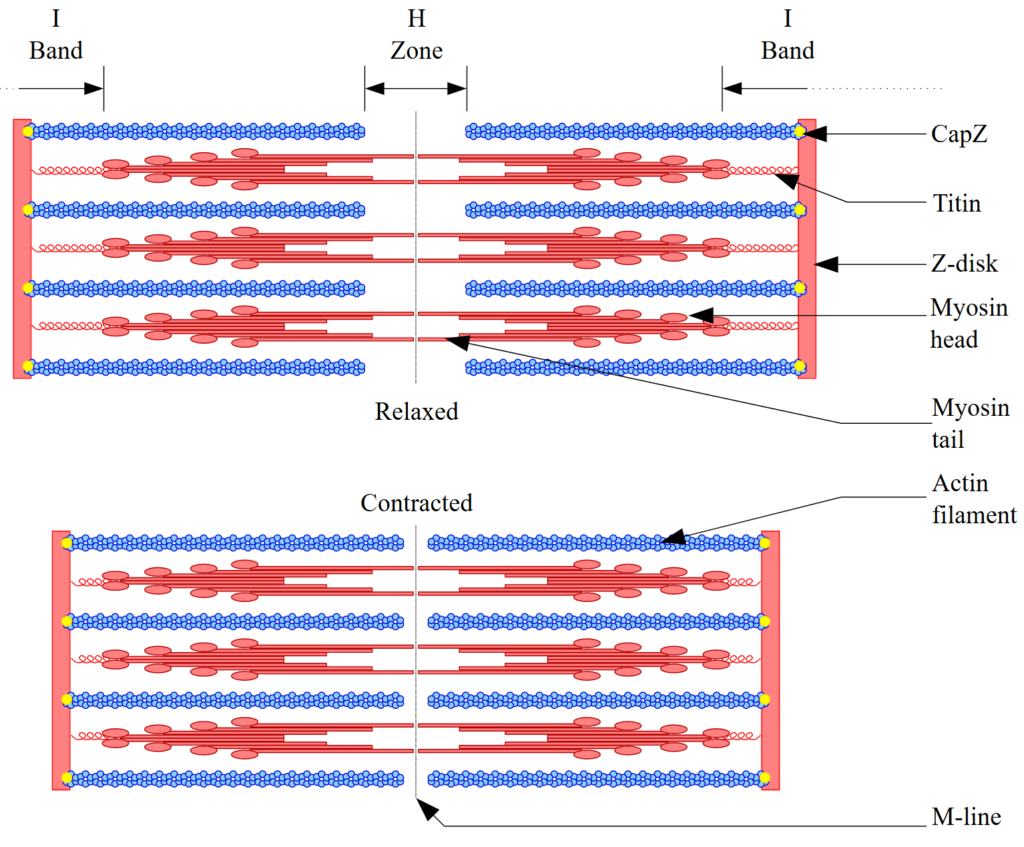
Credit: David Richfield under GNU Free Documentation License
Time for the biochemistry, you may want to hold your hat. The signals that will ultimately result in contraction of the skeletal muscles originate from the central nervous system, generally the brain, though in the case of reflexes it can also originate straight from the spinal cord since time is of the essence – this saves time both on travel and processing and can result in movements crucial in the odds of survival of the organism. A neurotransmitter then conveys the signal across a short synapse between a motor neuron and the muscle fibre by binding onto specific receptors; some of those technical terms will be elucidated in Chapter 6. The activation of the receptor liberates calcium cations (Ca2+) and to a lesser extent potassium cations (K+) thus causing an immediate reversal in polarity of the cell membrane (of the muscle fibre) called sarcolemma. This spike in positive charge within the cell is called depolarization – the technical definition is actually the internal charge becoming less negative, not necessarily becoming positive – and creates an action potential travelling along the cell, which ends up by activating a series of receptors that once again result in the release of Ca2+. Some of those calcium ions bind to a protein called Troponin C that, like actin, is part of the thin filament thus kicking-off the “cross-bridge cycle”.
I will try to summarize the sequence of this molecular cycle and, to compensate for this, I will include the Wikipedia entry to Muscle Contraction at the end of this chapter so you can read it either now or later in your spare time. You may recall from S1 Section 4.a that proteins folds in particular ways which allows them to perform particular functions and strongly bind to specific sites. In this case a cross-bridge consists in protruding parts of the myosin proteins, within the thick filament, with distinct binding sites for the ATP energy macromolecule and for actin, part of the thin filament. And here is the master stroke of nature, it is fittingly called “power stroke”: when ATP gets hydrolysed, some of the freed up chemical energy is transferred to the myosin and, courtesy of intramolecular forces, this permits a change in spatial orientation of the myosin head attached to the actin and by extension to the thin filament. This creates a pull or sliding of the two filaments in opposite direction. Then a new ATP macromolecule arrives and the head detaches before another cycle starts. Clever.
Distance of the sliding? About 10nm only but then the sarcomere density is about 250 per millimetre so we are looking at an aggregate contraction of over 2.5 micrometre per millimetre of muscle fibre and multiple contractions per second; once again volume does matter.
When it comes to relaxation, that state is achieved relatively simply by removing the concentration of Ca2+. The muscle is then able to contract again. The entire process, starting from depolarization of the muscle membrane is called excitation-contraction coupling (“ECC”).
Finally, time to zoom out. Repetitions of sarcomeres form myofibrils with diameters of 1 to 2 microns, a sub-unit or organelle of muscle fibre cells. In turn, the muscle fibres are bundled into fascicles surrounded by connective tissue, and the aggregate of those fascicles ultimately forms a specific muscle working in conjunction with other nearby muscles in what is functionally termed a muscle group such as the abdominals.
A discussion of muscles would not be complete without mentioning the three types of muscle fibres:
- Type I fibres contract slowly and relies on ATP produced through aerobic respiration, i.e. relying on oxygen and glucose.
- Type IIA also primarily rely on aerobic respiration but they do contract much faster than Type I and tire more quickly. The proper concept is actually muscle fatigue and it technically means the muscles’ ability to generate force is diminished, whether this is caused by a decline in the capacity of muscles to contract or neural fatigue.
- Type IIX also contract quickly but they do rely on an anaerobic pathway and thus fatigue even faster than Type IIA.
c) The human skeleton and locomotion
When I mentioned muscle contraction, you probably pictured a shortening of the muscle. That’s a mistake everybody makes initially; muscle contraction translates into tension and may or may not involve a change in muscle length. For instance, if you hold a cup of tea in your hand and stay still there is no muscle movement yet you are undoubtedly generating some force. Tension is essentially the force deployed by a muscle and is a function of the frequency of the action potential, meaning the number of times the neurons have fired to call for a muscle contraction, and of the number of fibres that are being called upon to contract. The mind boggles just thinking about the number of neurons firing taking place for the slightest of our moves…
When tension is created without involving a change in muscle length, this is called an isometric contraction (“iso” means same and metric conveys the idea of distance or movement). When the muscle shortens, we have a concentric contraction, think biceps curl or hamstring shortening causing your leg to move back when you walk or run. When the muscle lengthens, we have an eccentric contraction, this is your triceps extension and the phase your hamstring goes through before your foot hits the ground. If the muscle length varies and the tension doesn’t, then the contraction is isotonic.
All our movements rely on a combination of tension and changes in muscle length but, on their own, they would not result in locomotion. For this to happen, the muscle fibre needs to be connected to the bones of our skeleton, this connective tissue is known as tendons or sinew and is made of collagen, a structural protein. The bones themselves are connected via ligaments and sometimes cartilage is present between load-bearing bones where it acts as a shock absorber and reduces friction.
Consequently, proper locomotion not only requires highly coordinated muscle contractions managed by the central nervous system but it also calls for stability in the whole skeletal edifice, which is made possible via proprioception, the sensing of our own body position, the force and direction of our movements.
When it comes to the design of the skeleton and muscles, we have to defer to evolution and environment plays a crucial role there. For instance, aquatic animals are not heavily subjected to gravity so the frame of their skeleton need not be as strong as a land animals and their morphology would need to be optimized for the friction or resistance created by water, not something that is dramatically important for us humans surrounded by a gaseous atmosphere.
Time to have a quick sightseeing tour of the human skeleton. The official number of bones in adults is 206 though we are born with closer to 270 and some of them fuse during development. The key structural part is the axial component anchored by the vertebral column to which the rib cage and skull are attached. The hip bone provides the transition to the lower limbs attached at the hip joints; the main bones there are, from top to ground, the femur, the kneecap (patella), the tibia and fibula duo, the tarsals, metatarsals and phalanges of our toes. As for the upper limbs they are attached at the shoulder girdle consisting of the clavicles and scapulae; the main bones of the arms consist of the humerus, the ulna and radius duo, the carpals, metacarpals and the phalanges of our fingers. Voila.
d) Role and composition of the skin
Fortunately it seems, our musculoskeletal system is enhanced by wrap-around tissues, the external layers being formed by our skin and the rest of the integumentary system it is part of. As one would expect, the role of the skin is not simply aesthetic, it fulfils various functions that either provide a survival advantage or information to the organism.
The latter, the provision of information, is made possible by the embedding of sensory receptors detecting pressure, heat and cold, and the state of the skin itself in case of injury. The topic of touch will be covered properly in Chapter 9 so in this section I will focus on the other aspects that are all survival-oriented.
Arguably the most notable aspect of skin is its relative toughness compared to other tissues inside our body, this is even more obvious in other animals such as a crocodile. In effect, skin acts like a barrier, one that plays a role in both directions. It prevents intrusions, be it by pathogens and from shocks with the outside environment and it avoids fluids entering and escaping the organism, water in particular. In that sense, it can be thought of as a biological membrane except we are not dealing with mono or bi-layers of lipids but with stacked cells forming what is called the epidermis, the outermost part of the skin.
This organ is also actively involved in regulating the temperature of our body and does achieve this feat mainly via three different mechanisms. The first is the trapping of air within the hair, which is called fur when it takes a very dense appearance in some animal species. If you have read Series 1, you may recall the S1 Section 9.d on thermal conduction and convection, yet even if you have not, the physics can be grasped from a high level. What happens on an ongoing basis is that heat, energy in transfer, is leaving our body by radiation and conduction – when we touch a cold object for example. This creates a thin layer of warm air adjacent to our skin that gets replaced by convection, i.e. the warm air moves away because it is lighter than the ambient air. The partial trapping of this thin warm layer by our hair significantly decreases the loss of heat by convection and the effect is not negligible as the wind chill effect can attest to. The removal of the warm layer accelerated by wind boosts the rate of heat transfer and is perceived as a more pronounced cold by the thermal receptors in our skin.
The second mechanism for thermal regulation is the expansion or constriction of blood vessels which affects the throughput of fluid, thereby increasing or decreasing the rate of heat loss in the areas where blood flows near the skin and therefore close to the external environment.
The third mechanism bears the name of sweating and relies on the cooling effect of fluid evaporation. The eccrine glands in the skin secret a liquid known as perspiration or sweat on the outer surface of the skin. Within that liquid which is now exposed to the hot ambient air and therefore to gas molecules with high kinetic energy, some of the molecules with the highest kinetic energy will benefit from a transfer of energy from gas molecules and ultimately unbind themselves from the rest of the liquid molecules to join the higher energy world of gases. By definition, this vaporization first affects the liquid molecules with the highest energy so when they leave the liquid system, the average energy contained within decreases. That’s the cooling effect and it is much less pronounced when the atmosphere is already charged in moisture as it slows down evaporation rate, one of the reasons why training outdoors in tropical climates is quite challenging. You may want to read S1 Section 2.e on states of matter and phase transition to make better sense of this phenomenon.
The same way the skeleton performs secondary functions, the skin has been put to good use by our organism. Unlike some other animal species, we do not use it for camouflage, or at least not anymore, but we do use it for storage, mostly of lipids and water. Not to be mistaken with the adipose tissue known as subcutaneous fat located in a layer below the skin called the hypodermis which works rather well as padding to absorb shocks and, of course, as energy storage.
As mentioned earlier in this section, the skin is part of a broader set of layers and organs forming the integumentary system. The hypodermis belongs to that system together with elements such as glands, our hair and our nails, while the sensory apparatuses with receptors in the skin are grounded in the lower layers. A lot of the functions are one and the same as those listed for the skin and the distinction between the skin and other layers can be somewhat arbitrary from that perspective.
e) Trivia – Animal flight
We can walk, run, climb trees, jump and even swim. There is one thing we can’t do however; it is to be airborne for several seconds without external equipment and surviving to tell the tale. Not so for several animal species that have evolved aerial locomotion, be it unpowered in the case of gliding or powered in the case of flight. In this section we will direct our curiosity towards animal flight in birds and insects, noting that it has evolved a few more times, including in bats and the flying reptiles called pterosaurs, a species now instinct for probably over 60 million years.
The mechanics of flights do get actually quite tricky in the sense that a lot has to do with aerodynamics, that is the forces arising depending on the airflow created by the wing strokes. Therefore, I will try to provide a mildly technical explanation and if the fancy takes you, I have included two links to Insect Flight and Bird Flight entries on Wikipedia in the next section.
Wings in insects are part of the exoskeleton and can be linked to muscles in the case of direct flight and to other parts of the thorax in what is called indirect flight when they are made to move due to muscles deforming the shape of the thorax. The wing stroke that makes those organisms airborne can be broken down into two phases: the down stroke which produces the uplift in the air and the upstroke which brings the wings back to the starting position for the next downstroke. It goes without saying that on the way up the interaction of the wing surface with the air is not symmetrical to that on the way down, otherwise this would project insects back down towards the ground, a case of zero-sum forces. Instead, as the wing is about to move upward it rotates in a vertical position so as to minimize air friction during the vertical upstroke. Think about the way we swim freestyle, opening our hand after the catch to pull then push the water and project ourselves forward and, when our hand re-enters the water, we keep it horizontal with the surface to minimize resistance until the next catch and pull phase starts. Some insects have stroke frequencies that correspond one-on-one with action potentials but as the frequencies goes past a certain point, sometimes several hundred strokes per second, this is beyond the speed permitted by the nervous system which relies on neurotransmitter for the signal to bridge the synapses. In such cases, the biochemical reactions involved in muscle contractions and more specifically the removal of ions that start the relaxation phase are partially decoupled from the neuron action potentials.
The stroke mechanics of birds is actually quite similar to that of insects with the wing’s downstroke providing uplift and the upstroke seeking to minimize drag, however in the case of birds the wings tend to fold close to the body rather than pivot to a vertical position to achieve this.
The wings of birds have also evolved to provide a very significant surface which provides the ability to glide, thus reducing the required energy expenditure. All else being equal the air friction and gravity pull would result into birds slowly falling from the sky but they have learned to take advantage of convection in the form of rising columns of air, also known as thermals.
Minimizing energy expenditure is a huge driving factor in evolution and some bird species marooned on islands for thousands of generations may altogether lose their ability to fly, despite keeping their wings, if they lack the need for flight. This entails both the ability to feed themselves and avoid predators without being airborne. Think penguins or even ostriches.
f) Further reading (S2C3)
Suggested reads:
- The Body: A Guide for Occupants, by Bill Bryson (buy)
- Wikipedia on Muscle Contraction: https://en.wikipedia.org/wiki/Muscle_contraction
- Britannica on the Human Skeleton: https://www.britannica.com/science/human-skeleton
- Wikipedia on Insect Flight: https://en.wikipedia.org/wiki/Insect_flight
- Wikipedia on Bird Flight: https://en.wikipedia.org/wiki/Bird_flight
Disclaimer: the links to books are Amazon Affiliate links so if you click and then purchase one of them, this makes no difference to you but I will earn a small commission from Amazon. Thank you in advance.
Previous Chapter: The Digestive & Renal Systems
Next Chapter: The Endocrine & Reproductive Systems