>>> Click here to access this episode of the Syllab Podcast on Spotify <<<
a) Sound wave mechanics
The senses of hearing and olfaction are both sensitive to chemical compounds and vision relies on electromagnetic waves. Therefore, what is left for an organism to extract additional information from its environment and get an additional fitness advantage that can make all the difference in terms of survival and, ultimately, replication of the underlying genes?
The answer is mechanical stimuli such as pressure patterns and intensity. In the case of our auditory system, the nature of this mechanical stimulus is sound waves and it allows us and other animal species to detect, localize and even identify the source of those soundwaves with fairly high accuracy. Not only that but it also allows for intra-species exchange of information in the form of sound emission, and in more culturally advanced species, language. Think for a second of the advantage such a system provides as complement to the other senses: the ability to hear a predator or prey can make all the difference and making this information available to other members of a tribe to warn or attract them also confers an advantage at the group level and therefore, on average, for the genes present within the various organisms among the group.
We’ll look into the transduction process of sound waves, a type of mechanosensation, in the next section and after that we’ll seek to understand the process of auditory perception. However, before all of this, we need to spend some time on the specifics of a sound wave to understand the type of information it can convey. A sound is a mechanical wave propagating through various media such as gases, solids and liquids in the form of the compression and expansion of particles, therefore it carries a mix of kinetic energy in relation with its velocity and potential energy temporarily stored in the form of compression – it is slightly more complex for transverse waves with vertical oscillation but in fluids sounds are transmitted as longitudinal or compression waves.
As with electromagnetic waves, the physical attributes of a sound wave include its frequency (which is the reverse of its wavelength) and its direction of propagation. In addition, sound waves travel at different speeds and, even more importantly, they have different amplitudes or intensity, which determines the amount of pressure they exert on our mechanoreceptors and how loudly we hear them. And when I say we hear them, I mean how loud our central nervous system interpret them to be and somehow the phenomenon of qualia enables us to hear theses sounds in our mind. If the word qualia doesn’t ring a bell you should read through S2 Section 7.e.
Just to wrap up the physics aspect, the pressure is a function of the force applied by the vibration of the particles divided by the surface area upon which the pressure is applied and the way we measure sound level in decibels (dB) is as a logarithmic ratio between the intensity or pressure of the sound wave being measured and that of a reference intensity or pressure.
b) Transduction of sound in the ear
Now, onto the evolutionary marvel that is the ear. I’ll grant you it does not look like much from the outside but everything there is an engineering feat, even the outer ear that is sometimes called pinna in animals who can move it to improve focus depending on the direction of the sound waves. Indeed, the outer ear acts as a funnel channelling the waves into the ear canal in a focused fashion, and some more, as we’ll see in the next section.
At the end of the ear canal, the pressure of sound waves cause a thin membrane called the ear drum (or tympanic membrane) to vibrate. On the other side of the ear drum is the air-filled middle ear which comprises three tiny bones collectively called ossicles. It isn’t important though you may want to know those three bones are the smallest in the human body and called the stapes, malleus, and incus and more colloquially the stirrup, hammer, and anvil. The role of the middle ear includes transmitting those vibrations to the inner ear but, since that part is filled with liquid, it is also to regulate the loss of signal intensity which naturally occurs when pressure is transferred to a denser medium. The technical term for this is impedance matching and I include the link to the Wikipedia entry at the end of this chapter if this arises your curiosity.
In the inner ear, for purpose of explaining the auditory system, the structure of relevance is called the cochlea and within it the organ of Corti where the transduction of incoming pressure waves into electrical signals takes place. This feat is achieved by the pressure-driven movement of hair cells about 4-10 microns high that initiate a chemical cascade including the opening of ion channels and subsequent release of neurotransmitters which, after crossing a synapse, will generate an action potential at the level of the auditory nerves. I am blitzing through the concepts of chemical cascade, neurotransmitters, membrane depolarization and action potentials since we have now seen those play out and explained them in previous chapters, be it for our various senses and for other processes within animal organisms.
Interestingly, the hair cells are tonotopic, meaning they are specific to certain ranges of frequencies and since there is a variety of them, as a whole they are able to capture information regarding the mix of vibration frequencies they experience. To facilitate this, the hair cells are split into two categories, one called outer hair cells whose job it is to amplify vibrations of certain frequencies and the other called inner hair cells taking care of the transduction of signals at the frequency to which they are sensitive, in a graded rather than binary manner.
The tonotopic property of auditory hair cells is analogous to the variety of olfactory and gustatory chemoreceptors that bind to specific types of molecules, with more or less affinity, and provide data relating to the presence and concentration of molecules, thus helping our brain to identify chemical compounds and generate our experience of smell and flavours.
c) Auditory perception
The auditory nerve is directly embedded in the cochlea; hence it is also called cochlear nerve. It conveys information to the brainstem (in the cochlear nucleus to be precise) in the form of spatiotemporal patterns of neurons firing that reflect the amplitude and mix of frequencies of the sound waves initially collected by our outer ear.
Various partial processing steps occur first in the brain stem and then in areas of the mid-brain before signals reach the primary auditory cortex in the temporal lobe. This is where the final data parsing and treatment takes place, including the ability to untangle the various streams with different frequencies to isolate different sounds, a task that becomes difficult both as the mix increases in quantum, such as in white noise, and when separate streams have common frequencies. Ultimately this results in us hearing a sound but the job of ascribing meaning to those is handled by our language processing areas.
Not bad at all and yet, there is more. As alluded to in the initial section, our outer ears come in handy to derive further information. As was the case for our eyes and nasal cavities, our ears come in pair for a reason, which is to help ascertain the position of the source of the sound, and they are assisted in this by the asymmetrical shape of our outer ear – yes, in biology everything serves or did at one time serve a purpose.
- The horizontal angle of provenance of a sound, technically called the azimuth, is determined primarily based on the difference in the timing of receipt of the same sound between each ear since sound originating on our left side will first reach our left ear then the right one. Because our head is in between and creates a sound shadow, the difference in intensity between the sound collected in each hear acts as a second clue. To tell whether the sound is coming from a given angle in front of us or the same angle behind us, we rely on the shape of our outer ear which refracts pressure waves differently depending on the direction it comes from, thus yielding a slightly different acoustic pattern.
- Similarly, the shape of our outer ear and ensuing refraction patterns helps us discriminate between sounds coming from below or above us, i.e. pinpoint their provenance on a vertical plane. This cue is primarily extracted from high-frequency sounds whereas differences in timing used to establish azimuth are best extracted from low-frequency sounds.
- As for the distance parameter, also called near-far dimension, it is established based on two variables. The first one relies on prior experience of the typical intensity of a particular noise and thus, by comparing the intensity of the sound one just heard with its standard intensity at a given distance, it is possible to probabilistically infer how far away the noise source is. The second type of information is not completely reliable either since it supposes the presence of reflective surfaces. Assuming there is at least one, because the power of the reflected sound wave attenuates with the distance, the ratio between the original and reflected waves provides a cue to establish distance.
d) The vestibular sense
We’ll stay with the ear and more specifically the inner ear because this is the key peripheral location for the vestibular sense, that which helps us achieve balance and complex movement coordination. In S2 Section 3.a, we saw that the main functions of the musculoskeletal system could be grouped into four: movement, stability, organ protection, and form. Organ protection won’t be of concern here but the first two, movement and stability, are the result of complex muscle actions that need to be timed with respect to each other. This calls for several types of information such as limb position, hence the role of form in an organism and our nervous system mental map of it, the force being deployed by our muscles and experienced by each limb, as well as the position in space of our body as a whole and the nature of its movement through space.
Accordingly, the human species and other vertebrates have developed two sets of information gathering systems which, when integrated, enable us to perform complex tasks with our hands, feet and not fall every few seconds in the process. For limbs, the system is called proprioception and encompasses data about #1 the force exerted or load experienced by our muscles, #2 the way muscles are spatially translated and therefore limbs move, and #3 the position of the latter. We will look at proprioception in the next section.
For our body as a whole, the information can be broken down into the detection of its orientation and the linear acceleration it is experiencing; this is termed the vestibular sense and it is the topic we will pursue now. By the way, I think it is useful to remind ourselves that, as we saw in S1 Section 10.a, gravity is an acceleration and the level experienced on average on Earth is equal to 9.8 m/s2, also known as “1g”. This means even when we stand erect or sitting down our senses will be able to work this out by detecting a vertical acceleration towards the floor and no acceleration on the horizontal plane.
Besides the cochlea used to transduce pressure waves into electrical signals carried by the nerve, the human inner ear harbours a set of structures called semicircular ducts and a pair of otolithic organs called the utricle and the saccule. Figure 5 below provides a useful visual illustration of the inner ear in which all of these are depicted.
Figure 5: Representation of the human inner ear
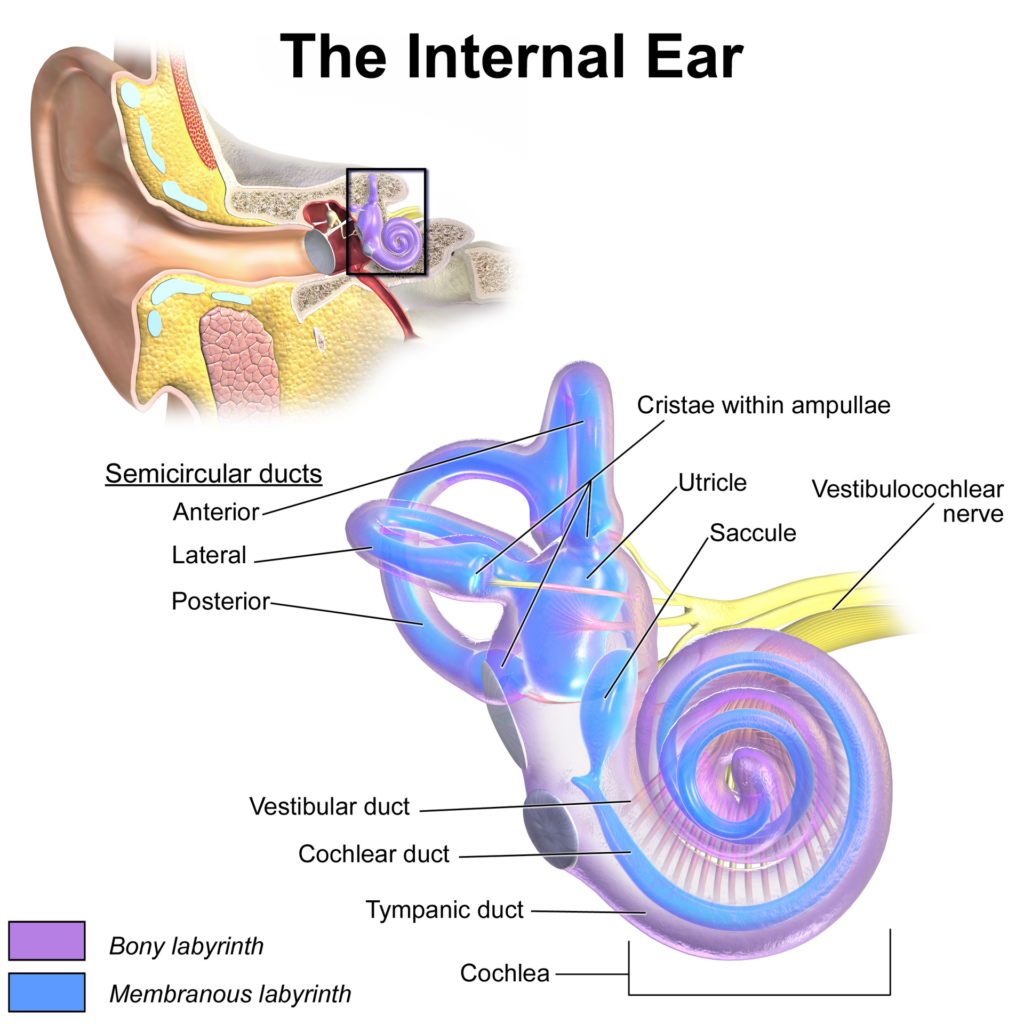
Credit: Medical gallery of Blausen Medical 2014 (CC BY 3.0)
The semi-circular canals come as a trio in each ear, each roughly perpendicular to the other, which should be expected if we are to derive 3-dimensional information. These canals rely on the movement of fluid within them to inform left-right rotation of the head on the horizontal plane, the vertical rotation in front or behind us (sagittal plane) and the vertical rotation on our right or left side (frontal plane). In order, these tasks are performed in the horizontal, anterior and posterior semi-circular canals, respectively. The information is gathered by hair cells that detect the acceleration of the fluid in the canals and since we have two ears, one on each side of the head, the spatial set up of the vestibular system is such that it is possible for the brain to work out the direction of the rotation on all three planes.
If the canals detect rotational movements, it is the role of the otoliths to capture linear acceleration-related data. The point is primarily to detect tilts of the head that announce or could lead to a fall and try to prevent it through a series of muscular actions. Once more, our organism relies on hair cells and the pressure applied on them by a membrane, with the detected intensity of pressure changing depending on the acceleration experienced in all three planes. By integrating with other cues, including receptors in the neck and inputs from the visual system, our central nervous system is able to differentiate between accelerations impacting the entire body and those only applying to the head, as when we nod.
How are the signals originating in the hair cells transduced along the nerves and up to the brain? Chemical cascades again, no surprise there if you have read the previous chapters and even section b) above. In this case, the movement of part of the hair cells causes the opening of ion channels and an influx of potassium cations resulting in the depolarization of the hair cell. In turn, this opens calcium channels and the liberation of neurotransmitters that will ultimately generate action potentials within the neurons of the vestibular nerve, which will join the cochlear nerve to convey signals to the brainstem for partial processing and onward transmission to other parts of the brain.
Before moving on to proprioception, it is worth mentioning a couple of reflexes that respond immediately to vestibular stimuli. The first is called vestibulospinal reflex and triggers micro-adjustments in posture via our skeletal muscles to ensure we maintain posture and therefore balance. The second is called vestibulo-ocular reflex and, like its name suggests, it involves movements of the eyes to compensate movements of the head and preserve as much stability in the visual field as possible. Thus, if you look at a tree and turn your head towards the right while maintaining your gaze on this tree, this is made possible by a corresponding movement of your eyes in the opposite direction, towards the left.
In our modern world, sometimes our central nervous system struggles to reconcile contradictory or unusual combination of stimuli, and this we experience as nausea or vertigo. For example, in a car cruising at a regular speed we will not experience horizontal acceleration and yet our vision tells us we are moving on the horizontal plane. And in said car if you start reading a book your visual system suggests no motion whereas your vestibular system senses some small rocking and rolling depending on the quality of the road and various turns. Same thing for seasickness so the worst think you can do on a ship is to look at stationary items below deck and instead you want your visual system to experience the motion first-hand to reduce the conflicts in stimuli your brain is exposed to.
e) Proprioception
Unlike receptors for the vestibular sense that are concentrated in the inner ears, proprioceptors are located throughout our musculoskeletal tissues, be they tendons, joints or the muscles themselves. These in turn will, based on an underlying map of their respective “standard” location and default states of activation, permit our central nervous system to continuously update the mental representation we have of our body. As mentioned earlier in this chapter, there are three parameters our brain requires with regards to limbs and muscles: position, nature and velocity of movement, and muscle load.
Starting with movement, muscle spindles are stretch receptors embedded within the muscles themselves, they provide information with regard to changes in the length of the muscle, and over time about this rate of change so that both the nature of the muscle contraction or dilation and its velocity can be established. The muscle spindles are mechanoreceptors formed by bunches of muscle fibres that will open ion channels and cause a firing of the sensory neurons depending on the degree to which they are stretched.
The proprioceptors responsible for recording information in relation with muscle load, that is the resistance muscles are experiencing, are called Golgi tendon organs. These are located at the junction between tendons and muscles and are designed to detect states and changes in muscle tension. Similarly to muscle spindles, they are mechanoreceptors and the compression occasioned by the exertion of muscle tension opens ion channels that depolarize the axons of the sensory neurons and generate action potentials with the frequency of firing indicating the amount of force developed by the muscle.
The third and final type of proprioceptor is called joint receptor and it informs about the position of limbs by detecting joint rotation, a set of data points from which the relative angle of the bones on either side of the joint can be computed by our brain. Once more, these mechanoreceptors rely on the tried and tested ion channels and membrane depolarization to create signals travelling along the nerves.
When proprioceptive and vestibular information are integrated, we can accomplish complex tasks such as walking, writing or playing tennis. The last one I picked because it exemplifies the exacting balance and hand-eye coordination which is called for to time the proper movement so as to hit a fast-moving ball and send it over the net in a particular area, all this without falling.
If you recall the feedforward aspects of the visual system, we should fully expect this to play a role in the way we move, meaning there is always an anticipatory element at play. And so, at times, expectations do not match with the reality and can cause some temporary conflicts inside our mind. For example, as we step on a travelator, we take a couple of seconds to mentally adjust to the increased speed at which our surroundings move, as perceived by our visual system, and when we step off there is also a lag during which our surroundings seem to move faster than they actually are. Anybody who has run a long-distance race such as a marathon may also have experienced this end-of-travellator feeling when stopping just after the finishing line. This expectation is also at the origin of the “broken escalator” phenomenon, when the stationary nature of the escalator goes against our previous experience so that we almost feel like the escalator is moving backward when we step on it.
f) Trivia – Tropism
Plants may not need the same sense of balance as we have but still, they need to orientate themselves and grow or turn in certain directions in search for nutrients or sunlight. This is called tropism and the most common forms are in response to the presence of water, light, and chemicals, not forgetting the sensing of gravity informing which way a plant should grow its roots.
Phototropism is growth in response to light and although the answer is generally to grow towards the light source, it can also be away from it. The chemistry behind it is actually rather complex and involves hormone gradients triggering chemical cascades affecting the turgor pressure, the rigidity of the cell walls (weakening it on the side opposite to the light) and asymmetrical elongation of the cells.
The information about which way is up, just as for which way is down, permits gravitropism, the growth in response to gravity-related stimuli. Somewhat similarly to the vestibular system of our inner ear, gravity causes pressure which opens ion channels and impacts the distribution of the plant hormone called auxin, thus leading to differentials in the growth rate of cells, a form of directionality.
The mechanisms for the growth in response to the presence of chemicals (chemotropism) or water (hydrotropism) are similar in many ways to those described above except that in those cases the receptors are chemoreceptors rather than photoreceptors or mechanoreceptors.
g) Further reading (S2C9)
Suggested reads:
- Wikipedia on Sound: https://en.wikipedia.org/wiki/Sound
- Wikipedia on Impedance matching: https://en.wikipedia.org/wiki/Impedance_matching
- Wikipedia on Eye-hand coordination: https://en.wikipedia.org/wiki/Eye%E2%80%93hand_coordination
Previous Chapter: Olfaction & Taste
Next Chapter: Touch & Interoception